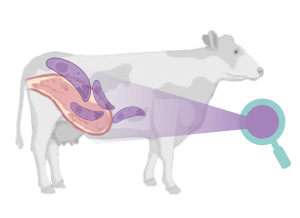
 La neosporosis bovina y la toxoplasmosis ovina son dos enfermedades parasitarias que causan importantes pérdidas económicas en el sector ganadero debido al fallo reproductivo que ocasionan.
La neosporosis bovina y la toxoplasmosis ovina son dos enfermedades parasitarias que causan importantes pérdidas económicas en el sector ganadero debido al fallo reproductivo que ocasionan.
 Estas enfermedades están causadas por Neospora caninum y Toxoplasma gondii, respectivamente, ambos son parásitos apicomplejos formadores de quistes estrechamente relacionados, y que comparten muchas características morfológicas y biológicas24,25.
Estas enfermedades están causadas por Neospora caninum y Toxoplasma gondii, respectivamente, ambos son parásitos apicomplejos formadores de quistes estrechamente relacionados, y que comparten muchas características morfológicas y biológicas24,25.
 La neosporosis bovina tiene una gran importancia en el ganado vacuno, ya que es una de las principales causas de aborto a nivel mundial, originando graves pérdidas económicas. El aborto es el principal signo clínico de la neosporosis bovina.
La neosporosis bovina tiene una gran importancia en el ganado vacuno, ya que es una de las principales causas de aborto a nivel mundial, originando graves pérdidas económicas. El aborto es el principal signo clínico de la neosporosis bovina.
 Los fetos de las madres infectadas por N. caninum pueden morir en el útero y ser reabsorbidos, momificados, sufrir autolisis o nacer muertos.
Los fetos de las madres infectadas por N. caninum pueden morir en el útero y ser reabsorbidos, momificados, sufrir autolisis o nacer muertos.
Las vacas de cualquier edad pueden abortar desde los 3 meses de gestación hasta su término, observándose la mayoría de los abortos a los 5-7 meses de gestación21.
Si la infección ocurre después de la mitad de la gestación pueden nacer terneros vivos con signos clínicos o clínicamente normales, pero persistentemente infectados.
 El fallo reproductivo puede producirse también en otros rumiantes como ovejas, cabras y ciervos21,26,27.
El fallo reproductivo puede producirse también en otros rumiantes como ovejas, cabras y ciervos21,26,27.
 En la actualidad, no existen vacunas ni tratamientos eficaces contra la infección por N. caninum28,29 y las opciones de control se basan en el diagnóstico y en medidas de bioseguridad y manejo21. En la actualidad, no existen vacunas ni tratamientos eficaces contra la infección por N. caninum28,29 y las opciones de control se basan en el diagnóstico y en medidas de bioseguridad y manejo21. |
 Por otro lado, la toxoplasmosis se considera una de las principales causas de pérdidas reproductivas en ovejas y cabras en todo el mundo22,30.
Por otro lado, la toxoplasmosis se considera una de las principales causas de pérdidas reproductivas en ovejas y cabras en todo el mundo22,30.
La infección durante la gestación suele provocar
la muerte del feto (infección al principio de la gestación); el nacimiento de corderos muertos o débiles, a veces acompañados de un feto de tamaño más pequeño y momificado (infección a mitad de gestación); o el nacimiento de corderos sanos, pero infectados congénitamente (infecciones durante el último tercio de la gestación)31. Es responsable del 10-23% de los abortos ovinos en Europa y EE.UU.32 y podría ser responsable de entre 680.000 y 1.360.000 abortos anuales en la UE. La toxoplasmosis es además una importante zoonosis y los pequeños rumiantes pueden desempeñar un papel importante en su transmisión a los humanos33. El control de la toxoplasmosis ovina se basa principalmente en la prevención de su transmisión horizontal y en el establecimiento de un programa de vacunación con una cepa viva atenuada S48 (Ovilis Toxovax®). ¿QUÉ TIENEN EN COMÚN N. caninum y T. gondii? N. caninum y T. gondii comparten muchas características en sus ciclos biológicos, sin embargo, ambos difieren en su rango de hospedadores, el potencial zoonósico (demostrado sólo para T. gondii) y la relevancia de las vías de transmisión horizontal y vertical en el mantenimiento de las infecciones en la naturaleza21,34. En ambos casos, los hospedadores definitivos son los únicos capaces...
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Phasellus non massa sit amet risus commodo feugiat. Quisque sodales turpis sed felis scelerisque, et luctus sapien facilisis. Integer nec urna libero. Sed vehicula venenatis lorem. Aenean fringilla dui non sapien pulvinar, sed tincidunt turpis tempus. Cras non nulla velit.
Regístrate y accede a los mejores artículos sobre rumiantes en rumiNews.
Regístrate en rumiNews para desbloquear este artículo y obtener más beneficios como:
Iniciar Sesión
Acceso a la
revista digital
Videos y podcasts exclusivos
Newsletter con las
últimas novedades
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Phasellus non massa sit amet risus commodo feugiat. Quisque sodales turpis sed felis scelerisque, et luctus sapien facilisis. Integer nec urna libero. Sed vehicula venenatis lorem. Aenean fringilla dui non sapien pulvinar, sed tincidunt turpis tempus. Cras non nulla velit.





 Latinoamérica
Latinoamérica
 Europa
Europa

































